Article Text

Abstract
Objective: To investigate the detailed anatomy of the terminal crest (crista terminalis) and its junctional regions with the pectinate muscles and intercaval area to provide the yardstick for structural normality.
Design: 97 human necropsy hearts were studied from patients who were not known to have medical histories of atrial arrhythmias. The dimensions of the terminal crest were measured in width and thickness from epicardium to endocardium, at the four points known to be chosen as sites of ablation.
Results: The pectinate muscles originating from the crest and extending along the wall of the appendage towards the vestibule of the tricuspid valve had a non-uniform trabecular pattern in 80% of hearts. Fine structure of the terminal crest studied using light and scanning electron microscopy consisted of much thicker and more numerous fibrous sheaths of endomysium with increasing age of the patient. 36 specimens of 45 (80%) specimens studied by electron microscopy had a predominantly uniform longitudinal arrangement of myocardial fibres within the terminal crest. In contrast, in all specimens, the junctional areas of the terminal crest with the pectinate muscles and with the intercaval area had crossing and non-uniform architecture of myofibres.
Conclusions: The normal anatomy of the muscle fibres and connective tissue in the junctional area of the terminal crest/pectinate muscles and terminal crest/intercaval bundle favours non-uniform anisotropic properties.
- anisotropy
- atrium
- catheter ablation
- electrophysiology
Statistics from Altmetric.com
The terminal crest is a significant structure in several forms of atrial tachyarrhyhmias and, occasionally, it is the target for radiofrequency catheter procedures. In common atrial flutter, the terminal crest acts as a natural barrier to conduction.1 Such block in conduction across the crest in patients with common atrial flutter2 seems to be determined functionally rather than anatomically, since transverse conduction is known to be preserved in some patients. Studies using intracardiac echocardiography have shown that two thirds of focal right atrial tachycardias seen in the absence of structural heart disease arise along the terminal crest.3 Ablation targeting the crest has also been used in patients with inappropriate sinus tachycardia.4 Despite its relevance to clinical and experimental studies, the architecture of the terminal crest in the human heart has not been studied in detail. Using an extensive collection of human hearts, we have determined the width and thickness of the terminal crest at four points and charted the variations in the trabecular arrangement of the pectinate muscles that originate from the terminal crest. In addition, in selected cases, we analysed the fine structure of the crest at these four points, elucidating the changes in the orientation of the bundles of fibres and the arrangement of the connective tissue. The latter structural features may explain why the crest can function as a functional barrier in patients with isthmic atrial flutter.
METHODS
We examined 97 randomly selected hearts from patients with unknown medical histories. All specimens were retained in accordance with guidelines for collection of human organs for research in Spain and Portugal. They were from 75 male and 22 female patients. The patients ranged in age from 8–91 years, with a mean (SD) of 49 (20) years. The weight of the hearts varied between 152–512 g, with a mean of 403 (60) g. Of the 97 specimens, 8 (9%) exhibited structural heart disease, consisting of ventricular hypertrophy in 3 (3%), and coronary arterial disease in 5 (6%). The intact hearts were fixed for at least two weeks by immersion in 10% formalin. The right atrium was opened through an incision in its intercaval wall, approximately parallel to the terminal groove, and then by an incision in the anterior wall of its appendage that extended into the superior vena cava and into the anterolateral side of the tricuspid annulus. These approaches allowed an adequate visualisation of the terminal crest and the muscular trabeculations originating from it—the so called pectinate muscles. We based our gross descriptions on the attitudinal orientation recommended by Cosio and colleagues on behalf of the Cardiac Nomenclature Study Group.5 Using calipers, we measured the crest at the following four points, noting its width and its thickness from epicardium to endocardium (fig 1A):
the origin of the crest from the superior rim of the oval fossa
at the level of the “precaval bundle”, which corresponded epicardially to the crest of the right atrial appendage
at the junction of the superior vena cava with the posterior atrial wall
halfway between the mouths of the superior and inferior venae cava.

(A) Opened right atrium in simulated right anterior oblique view to show the horseshoe shaped terminal crest (double dotted line). The crest arches anterior to the orifice of the superior vena cava (SCV) and extends to the area of the anterior interatrial groove. Inferiorly, the crest turns in beneath the orifice of the inferior vena cava (ICV), breaking up into a series of trabeculations in the area of the atrial wall known as the inferior isthmus (brace). Numbers 1 to 4 mark the points at which the thickness and width were measured. (B) The right atrium opened along the incisions marked by blue asterisks and the lateral wall deflected so as to display the crest (TC) on the endocardial surface separating the smooth walled venous component from the trabeculated appendage (RAA). The white asterisks are the cut surfaces of the crest. Note the ramifications in the inferior isthmus (brace). (C) Anterior aspect between the superior vena cava (SCV) and right appendage. The “septum spurium” (SS) is the most prominent anterior pectinate muscle arising from the crest (white asterisk). OF, oval fossa; TV, tricuspid valve; CS, coronary sinus.
At the same time, we studied the pattern of the pectinate muscles originating from the crest and extending along the wall of the appendage towards the vestibule of the tricuspid valve.
Scanning electron microscopy and histology
We made cross and longitudinal sections of the crest at the four levels identified above in 25 hearts selected to represent the age range. The small blocks of myocardium were removed and processed according to a modification of the cell maceration method.6 They were then dehydrated in graded acetone and dried to the critical point using carbon dioxide. The dried specimens were mounted on metal stubs, coated with gold, and observed under a Jeol JSM 5600 scanning electron microscope. Another 10 non-macerated specimens were prepared in similar fashion. The blocks from an additional 10 specimens were processed for conventional light microscopic study. Deparaffinised sections were stained with Masson's trichrome technique and the picrosirius red method. Sister histological sections were deparaffinised in xylene for 30 minutes until they were transparent, then dried at room temperature and glued to aluminium stubs, sputter coated with gold, and examined under the scanning electron microscope. From the sections and blocks, we evaluated the direction of myofibres along the length of the terminal crest. Using microscopic criteria, we described uniformity when there was a parallel arrangement of myofibres and non-uniformity when the myofibres crossed each other or were irregularly oriented. We also assessed qualitatively the changes in the fibrous matrix with the age of the patient.
Statistical study
Results are expressed as mean (SD). Quantitative data were compared using a Student's unpaired t test. Quantitative variables were correlated by linear regression analysis. A probability value of p < 0.05 was considered to be significant.
RESULTS
Inspection from inside the right atrium found that the terminal crest (crista terminalis) was basically a horseshoe shaped or C shaped structure (fig 1A). Anteriorly, it inserted into the anteromedial wall of the right atrium situated to the left of the orifice of the superior vena cava. Throughout its length, the crest formed the boundary between the smooth walled venous component of the atrium and the extensive appendage. Thus, the crest extended laterally and inferiorly, turning in beneath the orifice of the inferior vena cava to ramify as a series of trabeculations in the area of the atrial wall known as the inferior(or flutter) isthmus (fig 1A). In each heart, the origin of the crest at the interatrial groove was confluent with the origin of another important muscular fascicle, Bachmann's bundle, which extended into the left atrium. Table 1 lists the dimensions of the terminal crest at its origin (point 1) and the other three sites along its course. Comparisons between hearts from male and female patients showed significant differences in width (p < 0.001) and thickness (p < 0.01) only at point 4, which is located halfway between the caval orifices. However, the differences may be caused by the older age of the women in this study. In both groups, the crest decreased in width and thickness distal to point 1 but the differences were not significant. When traced laterally from its origin, at a site that corresponded epicardially with the crest of the right atrial appendage (point 2), the terminal crest gave rise to a series of relatively thick bundles, the anterior pectinate muscles, which fanned out anteriorly towards the smooth vestibule of the tricuspid valve. In 80 specimens (82%), one of these anterior pectinate muscles was particularly prominent, tracking along the superior part of the appendage and spreading out in an irregular pattern across the body of the appendage (fig 1A,B). Described as the “septum spurium” or “sagittal muscle bundle”, this muscular branch was 4.5 (1.2) mm in width (range 3–7 mm). This structure could not be identified as a distinct entity in the remaining 17 specimens (18%).
Width and thickness from epicardium to endocardium of the terminal crest at the four points corresponding to fig 1A, according to sex
When traced laterally, the crest arched to the right of the orifice of the superior vena cava, descending the right lateral free wall at a right angle relative to its precaval segment. The pectinate muscles originating from this lateral part of the crest took an oblique course towards the vestibule of the tricuspid valve. In the more distal segment of the crest, the pectinate muscles were more widely spaced. They fanned out into the posterior atrial wall towards the vestibule (fig 1A), with the crest itself losing its structure as an individual muscular bundle. The crest divided into a series of trabeculations with considerable variability in the inferior isthmus. Broadly, we recognised three patterns of ramification. In 68 of the specimens (70%), we traced thick muscle bundles from the distal crest to the vestibule of the tricuspid valve, with other thick bundles extending obliquely to the orifice of the coronary sinus (fig 2A). In a further 21 specimens (22%), thinner bundles radiated fan-like from the distal crest, extending in almost parallel fashion obliquely towards the vestibule of the tricuspid valve and the orifice of the coronary sinus (fig 2B). In the remaining eight hearts (8%), the muscular extensions extended longitudinally towards the vestibule (fig 2).
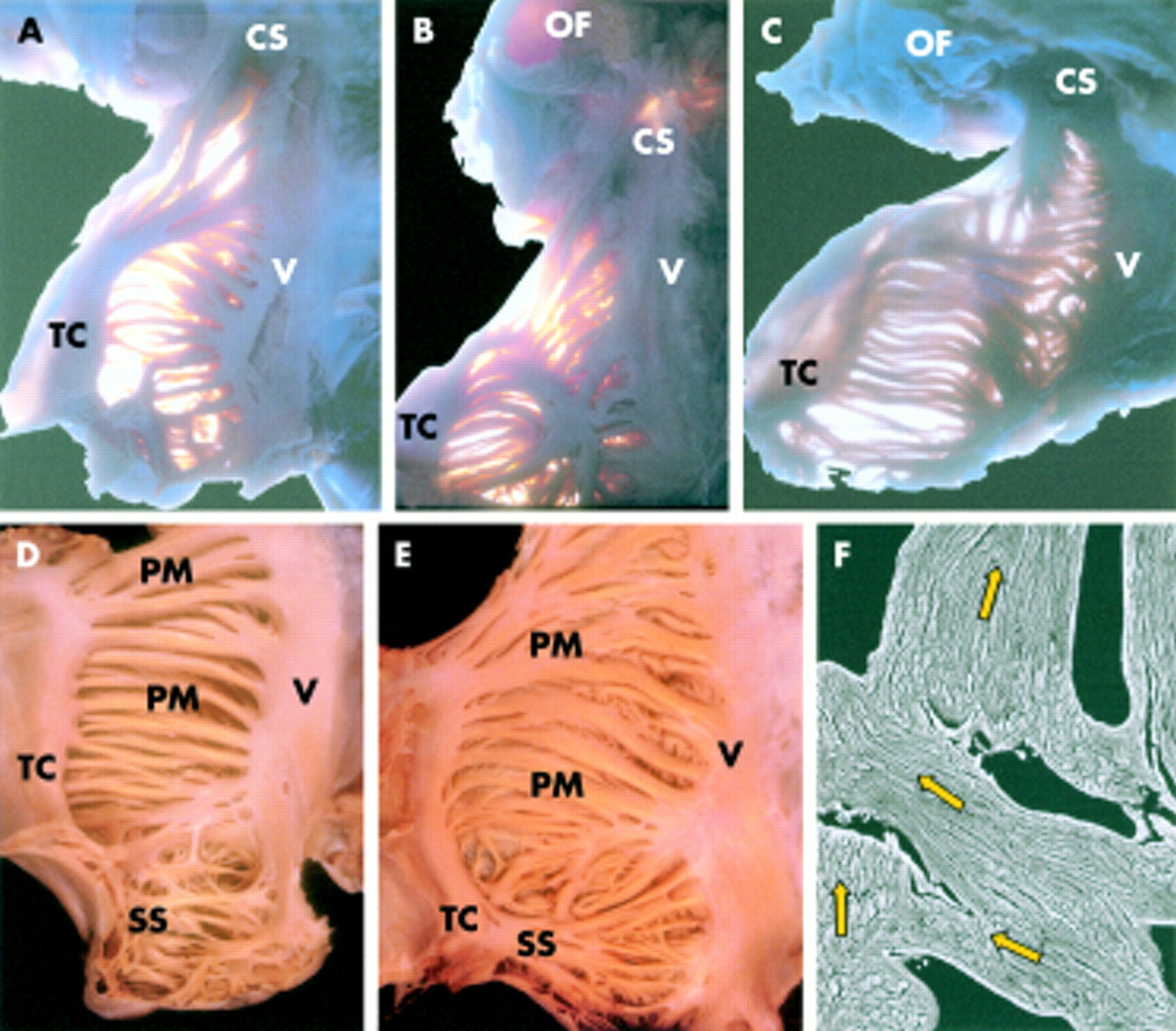
(A–C) Endocardial aspects of the lateral wall of the right atrium opened and displayed in the same orientation as fig 1B. Transillumination shows the variability (70%, 22%, and 8%) in branching of the terminal crest in the inferior isthmus. (D) Endocardial surface of the lateral wall has a criss-crossing trabecular architecture within the tip of the appendage whereas in the remaining wall the pectinate muscles have a uniform parallel alignment almost without crossovers between them. In contrast, the pectinate muscles in (E) have a non-uniform arrangement with abundant interlacing trabeculations between them. (F) Scanning electron micrograph of non-macerated specimen confirmed the irregular alignment (arrows) of the muscular myofibrils within the pectinate muscles. Original magnification (F) ×75. CS, coronary sinus; OF, oval fossa; PM, pectinate muscle; SS, septum spurium; TC, terminal crest; V, Vestibule of the right atrium.
The arrangement of muscle fibres in all pectinate muscles contained within the right atrial wall was diverse and differed from heart to heart regardless of age and sex. In all specimens, the pectinate muscles were particularly numerous within the tip of the anterior part the appendage, giving it a coral-like configuration, with a multitude of small meshes between them (fig 2D). In contrast, the pattern of the pectinate muscles along the lateral wall of the appendage was very uniform in 19 specimens (20%), with the muscular trabeculations extending from the terminal crest in almost parallel fashion, without appreciable crossovers but with areas between these bundles being paper thin (fig 2D). The remaining 78 specimens (80%) had abundant crossovers, with small interlacing trabeculations coursing between the individual pectinate muscles (fig 2E, F)
Microscopic and ultrastructural characteristics
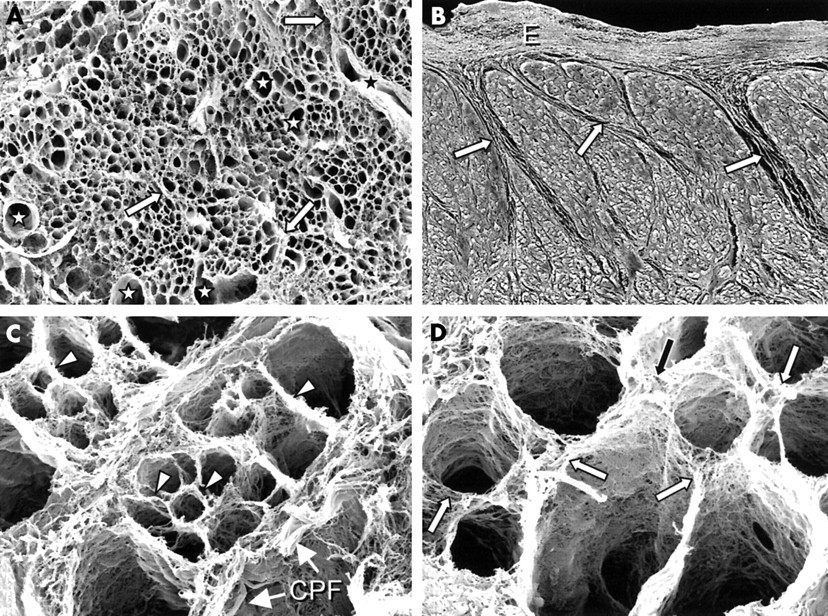
The scanning electron microscopic study showed a honeycomb organisation of the connective tissue skeleton of the crest in cross sections, interrupted only by thick collagenous septums of the perimysium and vascular spaces (fig 3A). Epicardially and endocardially, the crest was covered by a thick layer of tightly packed collagen fibres, which, at least in the periphery, were arranged circumferentially. The perimysial septums arose from either the epicardium or the endocardium and enveloped groups of myocytes. They were variable in number and spatial arrangement, running in different directions and never completely partitioning the crest (fig 3B). Coronary arteries and tendon-like collagen bundles coursed along these thick septums (fig 3C). The endomysium, as the final arborisation of the perimysium, supported and connected individual cells (fig 3C). The endomysial matrix consisted of very fine and smooth sheaths of collagen fibres arranged around each individual myofibre (fig 3D). It was connected to adjacent myocytes and interstitial microvessels by lateral struts (fig 3D).

(A) Scanning electron micrograph of a cross section through the terminal crest at point 3 after digestion with sodium hydroxide that removed the myocytes, leaving the fibrous matrix. At low power magnification, the presence of collagenous sheaths gives the terminal crest a honeycomb appearance, interrupted only by the presence of thick collagenous septums of the perimysium (arrows) and vascular spaces (stars). Original magnification ×120. (B) Scanning electron micrograph of non-macerated specimen shows the perimysial septums (arrows) arising from the endocardium (E). Original magnification ×65. Scanning electron microscopic images at high power magnification after digestion are shown in (C) and (D). (C) Collagen sheaths of the endomysium (arrowheads), which are oval or round, support and connect individual cells. Coiled perimysial fibres or tendon-like collagen bundles (CPF) course along the perimysial septum. Original magnification ×1600. (D) Endomysial sheaths are connected to adjacent myocytes by lateral struts (arrows) of variable width and length. Original magnification ×1500.
We noted great excess in the amount and tortuosity of the endomysial matrix around the cells of the sinus node, whereas we saw occasional perimysial septums between them (fig 4A), which increased in amount with advancing age (fig 4B). In the majority of hearts, 32 of the 45 studied (71%), the sinus nodal artery was located centrally within the node (fig 4A), although we noted an eccentric location in the remaining 29% of the cases. We were unable, however, to distinguish any particular organisation or spatial pattern of distribution of the endomysial struts within the node itself. Despite the presence of thick perimysial septums extending between the sinus node and atrial muscle, short tongues of nodal cells or transitional cells were seen to interdigitate with ordinary musculature of the terminal crest in non-uniform fashion. In 38 of the 45 specimens studied in this fashion (84%), the sinus node was found beneath the epicardium at the junction of the superior vena cava and the right atrial appendage. The distance between the epicardium and nodal tissue was 0.9 (0.7) mm (range 0–2.5 mm), the gap filled by fibroadipose tissue. In 7 of the 45 specimens studied with scanning electron microscopy (16%) there were small groups of nodal cells at the origin of the crest. The body of the sinus node was shown to be spindle shaped, which extended alongside the musculature of the crest subepicardially in lateral position, gradually passing through the musculature of the crest to become the nodal “tail” in the subendocardium lying halfway between the orifices of the caval veins. The distance from the right atrial endocardium to this tail varied around 1.5 (0.3) mm (range 1.1–1.9 mm).

(A) Scanning electron micrograph of the sinus node and nodal artery (NA) of a cross section through the terminal crest at point 2, after digestion of the nodal cells. The collagenous sheaths of endomysial formed a complex network in the gaps between the nodal cells, whereas occasional perimysial septums were seen between them. Original magnification ×65. (B) Scanning electron micrograph of non-macerated sinus node cells (N), which are encased in dense and coarse endomysial sheaths (arrows) in a specimen from a 75 year old. Original magnification ×600. (C) Scanning electron micrograph of a cross section through the crest, after digestion, from a specimen of 70 years shows a diffuse notable excess of endomysial sheaths (arrows) indicating focal interstitial reactive fibrosis. Vascular space (V) corresponds to a coronary vein. Original magnification ×360. (D) Scanning electron micrograph of non-macerated cross section through the body of the terminal crest shows mainly longitudinal fibres (H) with intermingling oblique (O) or lateral (L) fibres. Original magnification ×160.
In addition to the area of the sinus node, where extensive densely packed collagen was always observed, local differences in the pattern of accumulation of the collagenous endomysium was found throughout the length of the terminal crest, with variations according to age. With increasing age, the struts tethering myocytes to myocytes, or myocyte sheaths to endomysium, were much thicker and more numerous, indicating focal interstitial reactive fibrosis rather than replacement of myocytes in these areas (fig 4C). This feature, however, varied in intensity from area to area.
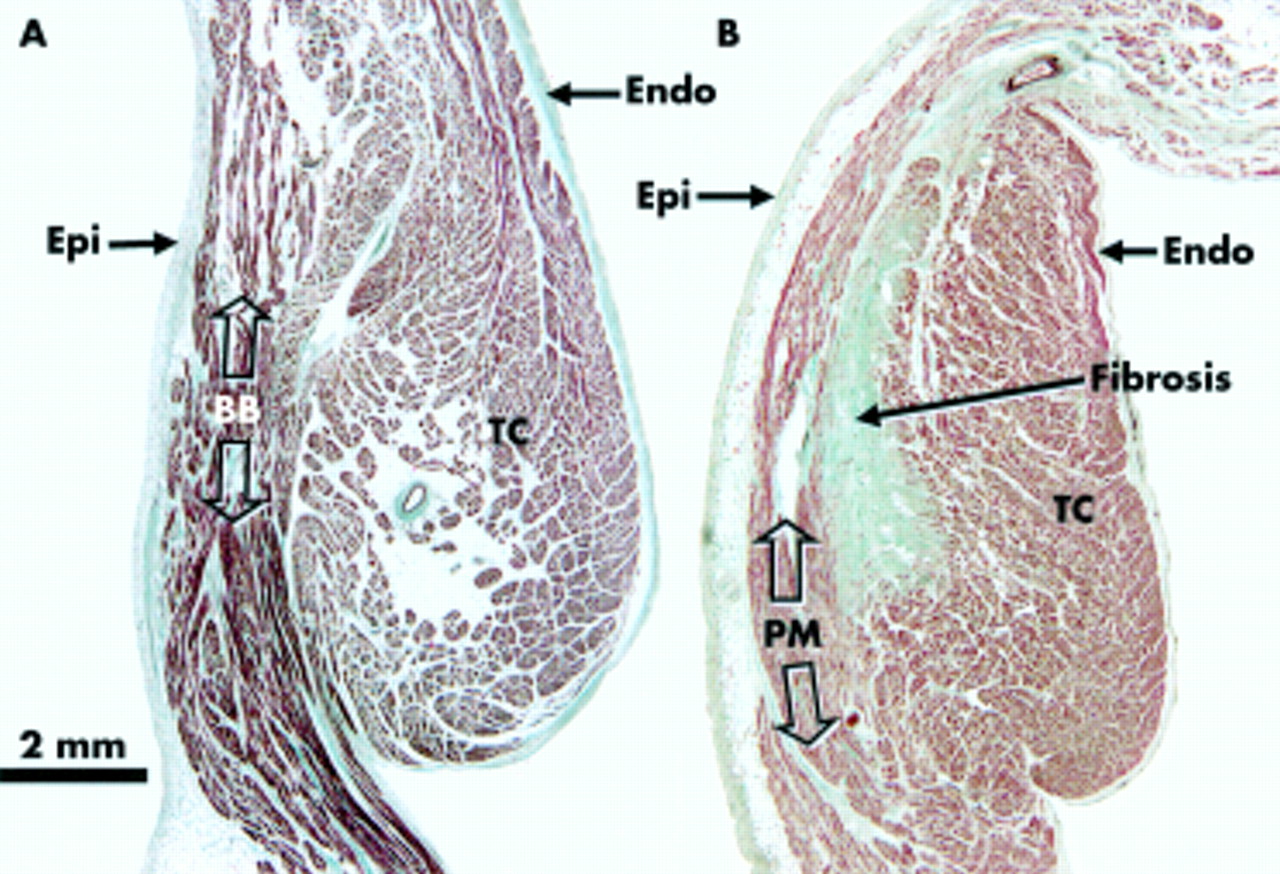
The myofibrillar architecture in the area between the origin of the crest itself and Bachmann's bundle was non-uniform in all specimens (fig 5A). In contrast, at the other three points sampled along the crest, most of the specimens (36 of 45 (80%)) had a predominantly uniform longitudinal arrangement within the crest. In the remaining nine specimens (20%), changes in orientation, or non-uniformity, of the fibres were also seen within the body of the crest (fig 4D). In all the hearts, at the margins of both subepicardial and subendocardial sides of the crest, the myofibres were irregularly oriented (fig 5B), intermingling horizontal with oblique or longitudinal fibres, and extending subsequently to reach the pectinate muscles or the wall of the intercaval region.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Cross section through the terminal crest (TC) at its origin (point 1 of fig 1A) showing its relation to the musculature of Bachmann's bundle (BB) in a 46 year old man. Note the change between the predominantly transverse muscle fibres of the terminal crest and the longitudinal fibres of Bachmann's bundle. (B) This section from a 76 year old woman taken at point 4 (fig 1A) shows a fibrotic area at the border between the terminal crest and the pectinate muscle (PM). Masson's trichrome stain. Endo, endocardium; Epi, epicardium.
DISCUSSION
The complexity of atrial geometry dictates that the spread of activation from the site of origin of the cardiac impulse will be restrained by naturally occurring barriers and orifices. It has been shown physiologically that the terminal crest provides a barrier to conduction transversely across it during typical atrial flutter.1,2 Mapping studies in animal models have shown the presence of such transverse conduction across the crest in normal hearts.7 Anisotropy within the crest, with a fast velocity of conduction in the longitudinal direction, and a low velocity in transverse direction, caused by the density of gap junctions at end to end connections, is also well established.8,9 Yamashita and colleagues7 were unable to find any systematisation of local effective refractory periods along the terminal crest. Rapid stimulation resulted in functional block of conduction at the lateral edge of the crest, as well as at its border zone with the pectinate muscles. These findings were supported by Schoels and colleagues10 using epicardial mapping in a canine model, while Schumacher and associates2 found that the crest provided the capacity for transverse conduction and that block along the crest in humans with atrial flutter was functional. Recently, Becker and colleagues,11 combining high resolution mapping with postmortem analyses in dogs, showed conduction block parallel to the long axis of the crest, and this phenomenon was common in two fifths of normal hearts studied. Despite these electrophysiological findings, the underlying mechanism causing the delay in conduction in atrial flutter or fibrillation remains unclear. Our study has shown that all “normal” hearts—that is, hearts obtained from patients supposedly without a clinical history of atrial tachyarrhythmias—contain a non-uniform muscular pattern in the margins of the terminal crest, at its junctions with the pectinate muscles and intercaval region. The posteromedial right atrium (sinus venosa region), corresponding to the intercaval region, has been identified as a functional line of block in patients with atrial flutter.12 Additionally in our study we found that the area between the origin of the crest itself and Bachmann's bundle, the anterosuperior part of the right atrium, had a non-uniform myofibrillar architecture in all specimens, providing some anatomical background to the reported electrophysiological studies.11 Universal irregularities at the margins suggest a pattern inherent to the terminal crest itself and may provide an explanation for the crest acting as a functional barrier in isthmic atrial flutter, rather than an anatomical barrier.
In concordance with the recent report by Waki and colleagues,13 our present study also showed that, in “normal” hearts, there is a non-uniform alignment prevailing within the terminal ramifications of the crest. This is the area of the atrial wall known as the inferior (or flutter) isthmus. We also showed that, in most specimens, the pectinate muscles running in the anterior wall of the appendage had abundant crossovers, with interlacing small trabeculations interconnecting the major muscular bundles. According to the model for anisotropic propagation in atrial muscle, such changes in orientation may, in part, account for discontinuities in the spread of the excitatory wavefront that may have a role in the genesis of atrial re-entrant arrhythmias. Non-uniform muscular patterns, therefore, can be considered the norm in the human right atrium, with the corollary that the potential for delayed conduction may also be a normal phenomenon. These are important anatomical observations that provide evidence concerning factors that can influence the route for conduction of impulses throughout the complex atrial geometry.
Furthermore, we found three distinct morphological patterns along the terminal crest. Firstly, there was a diffuse but notable excess in the amount and tortuosity of endomysial matrix around the cells of the sinus node, which increased in amount with advancing age. We could not distinguish any particular organisation or spatial pattern of struts running throughout the length of the node itself. Secondly, we found non-uniform alignment between the cells of the sinus node and ordinary atrial muscle because of the presence of thick perimysial septums, or short tongues of nodal or transitional cells, which interdigitated in an irregular pattern with the ordinary musculature of the terminal crest. Thirdly, we observed that, with advancing age, notable focal interstitial fibrosis developed around the ordinary atrial cells of the crest. The collagen struts have been described as being attached to the surface of the ordinary myocytes through branches that extend to the cell membrane at the Z bands.14 Our results show that the struts do not have preferential sites of insertion along the sarcomeric length. Spach and Dolber15 showed that the anisotropic electrical properties of human atrial muscle change with advancing age, with increasingly complex pathways of spread of propagation seen at a microscopic level. They concluded that the interactions between structure and cellular electrophysiological properties provide a basis for the understanding of various cardiac arrhythmias, such as the greater incidence of atrial tachyarrhythmias in older people. Our findings lend support to their hypothesis. Others, however, suggest that the predisposition of the crest for blocked conduction may be related to characteristic distribution of the channel proteins connexin 40, 43, and 45.9,16
Implications for radiofrequency ablation
Modification of the sinus node, closely related to the terminal crest, using radiofrequency has been established as a treatment for inappropriate sinus tachycardia.4 Because of the vicinity of the sinus node to the epicardial surface, with a mean distance of 0.9 (0.7) mm (range 0–2.5 mm) at the junction of the superior vena cava and the right atrial appendage, the sinus node is easily damaged if an epicardial approach is used or during cardiac surgery. Approaching from the endocardial surface, however, more radiofrequency energy would be necessary to ablate the sinus node, since the lesions need to penetrate through greater depth of the musculature of the terminal crest, which has a mean of 6.3 (1.5) mm (range 3–10 mm). Moreover, the centrally located nodal artery may provide a cooling effect, reducing the radiofrequency damage. These factors probably explain why radiofrequency current is less effective in suppressing inappropriate sinus node tachycardia than other right atrial tachycardias.17 In addition, when applied in these regions, radiofrequency current cannot only injure the myocardium of the terminal crest itself but can also induce venous occlusion at the superior venoatrial junction.18 From the anatomical standpoint, we suggest that positioning the ablating catheter at the site of the terminal crest halfway between the caval veins, where the tail of the sinus node is intramyocardial and located at a mean distance of 1.5 (0.3) mm (range 1.1–1.9 mm) from the endocardial surface, may be as effective in inducing block without the risk of causing venous occlusion.
Study limitations
An obvious limitation of the present study is that we have not been able to study hearts with documented atrial flutter or atrial fibrillation. Despite this, we submit that our investigation provides the necessary knowledge of normality on which to base a similar examination of hearts obtained from patients with atrial flutter. Although our method does not allow us to quantify the connective tissue or the angular changes in fibre arrangement at the boundaries of the terminal crest, it is useful for studying the spatial organisation of the myofibrils and collagen of the myocardium under normal conditions.
Acknowledgments
The authors thank Prof Dr Duarte Nuno Vieira and Prof Dra Maria Cristina de Mendonça, Instituto Nacional de Medicina Legal (Coimbra Bureau), Portugal, for allowing us to examine the specimens that formed the major part of the series. This project was supported by grant IPR00A037 from the Junta de Extremadura (DS-Q and VC), the British Heart Foundation (RHA), and the Royal Brompton and Harefield Hospital Charitable Fund and NHS Trust (SYH) and was facilitated by a travelling grant from Acciones Integradas.