Article Text

Statistics from Altmetric.com
In 1973 Chatterjee and colleagues, on the basis of clinical observations, suggested that a “marked improvement and even normalisation of left ventricular function can be expected in properly selected patients with the preinfarction syndrome or with chronic ischaemia and no previous infarct, following appropriate aortocoronary artery bypass surgery”.1 Five years later Diamond and associates were the first to use the word “hibernation” in the introduction to an experimental study on postextrasystolic potentiation in ischaemic dog myocardium.2 Somewhat vaguely, they concluded from the “sometimes dramatic improvement in segmental left ventricular function following coronary bypass surgery” that “ischaemic non-infarcted myocardium can exist in a state of function hibernation.” In the early 1980s, Rahimtoola systematically reviewed the results of coronary bypass surgery trials and identified patients with coronary artery disease in whom chronic left ventricular dysfunction improved on revascularisation.3 ,4 He then specified the term “hibernation” previously used by Diamond and associates, and proposed that the observed contractile dysfunction was not the result of an ongoing energetic deficit but an adaptive downregulation of contractile function to preserve myocardial integrity and viability.4 Thus the concept of myocardial hibernation challenged the traditional view of myocardial ischaemia which assumed that the extent of chronic contractile dysfunction reflected the amount of infarcted tissue. This new pathophysiological concept of hibernating myocardium was further popularised by Braunwald and Rutherford,5 who emphasised the need for its recognition and treatment through revascularisation.
When proposing the concept of hibernation, Rahimtoola reasonably assumed that the observed reduction of contractile function reflected a situation of reduced resting blood flow.4 Experimental studies indeed showed a proportionate reduction in regional myocardial blood flow and contractile function in response to graded reductions in coronary flow in dog hearts during acute6 and subacute ischaemia,7 as well as in pig hearts during sustained stress induced ischaemia.8 On the basis of such studies Ross introduced the concept of perfusion–contraction matching, and this was rapidly accepted as the basis of hibernating myocardium.9 However, in contrast to the clinical situation, all experimental studies showing perfusion–contraction matching were limited to observation periods of no more than five hours. Ross therefore proposed a distinction between “short term” hibernation, as observed in the experimental setting, and “chronic” hibernation, as seen in the clinical setting.
Perfusion–contraction matching of resting blood flow and function in short term hibernation is unequivocal (reviewed by Heusch10). Most studies in patients show a significant reduction in resting regional myocardial blood flow in dysfunctional areas classified as hibernating compared with normal areas remote from that site, with an average reduction in blood flow from baseline of 20–30% (see Heusch10). However, except in one study,11 wall motion scores were used and no absolute data on regional contractile function were provided. Thus the question about perfusion–contraction matching in chronically dysfunctional myocardium remains unresolved at present, and quantitative data on both regional myocardial blood flow and contractile function before and after revascularisation are required to answer it.
Metabolism of hibernating myocardium
Within the first five minutes of an acute coronary inflow reduction in anaesthetised pigs, coronary venous pH and lactate extraction were shown to be reduced and coronary venous Pco 2 to be increased, but these variables gradually returned towards control values during 180 minutes of continued moderate ischaemia.12 An early increase in myocardial lactate production followed by a decline to normal or near normal levels during sustained moderate ischaemia has been confirmed in several subsequent studies.13-18
In the in situ porcine model of short term hibernation, cardiac glycogen content was somewhat (though not significantly) reduced within the first five minutes of moderate ischaemia, but thereafter remained constant when ischaemia was prolonged to 90 minutes.14Glucose uptake was gradually increased within the first 20 minutes of moderate ischaemia in anaesthetised pigs19 and remained enhanced during 24 hours of coronary hypoperfusion,16 ,20as it did in pigs with chronic coronary stenosis and a persistent reduction in regional myocardial blood flow.21 ,22
In patients with a reduction in resting blood flow in dysfunctional myocardium, myocardial glucose utilisation did not differ from that in healthy volunteers,23 nor from utilisation in a remote reference region in the same patient,23 ,24 again showing that the ratio of metabolism to blood flow is increased in dysfunctional myocardium.
A decrease in ATP content over time was observed during continued myocardial ischaemia in isolated rabbit25 and piglet26 hearts, and in anaesthetised pigs13 ,14 ,27 and dogs.28 In contrast to the steady decline in ATP content, but in agreement with the attenuation of lactate production over time, myocardial creatine phosphate content was decreased immediately after the onset of ischaemia but gradually recovered over time towards control values,27 whereas regional myocardial blood flow and contractile function were persistently reduced.13 ,14 ,26-28 In chronically instrumented dogs, biopsy samples from the hypoperfused, dysfunctional myocardium revealed normal histology and high energy phosphate content.29 Sequential biopsy based measurements of ATP, creatine phosphate, creatine, and inorganic phosphate showed a decrease in the free energy change of ATP hydrolysis during early ischaemia, with subsequent recovery during continued ischaemia30 (fig1). Similarly in patients undergoing coronary artery bypass grafting (severe stenosis of the left anterior descending coronary artery), the ADP to ATP ratio, the energy charge, and the creatine phosphate content were within the normal range in the dysfunctional but viable (from histology) anterior myocardium.31 The most plausible explanation for the recovery of creatine phosphate content and free energy change of ATP hydrolysis is downregulation of contractile function—that is, energy demand.

Sequential biopsy based measurements of the free energy change of ATP hydrolysis. Empty circles are data from single animals; filled circles are mean values; error bars represent 1 SD. The free energy change of ATP hydrolysis is decreased within the first five minutes of ischaemia, but subsequently recovers towards control values when ischaemia is prolonged to 90 minutes. Reproduced from Martin C, et al. Cardiovasc Res 1998;39:318–26, with permission of the publisher.
Glycolytic ATP production and loss of adenine nucleotides25 may contribute to the restoration of an energetic steady state but probably play a relatively small role. In support of this view, pharmacological reduction of contractile function by intracoronary lidocaine prevented the decreases in ATP and creatine phosphate otherwise seen during a pronounced reduction in regional myocardial blood flow in anaesthetised pigs.32
Inotropic reserve in hibernating myocardium
Although baseline contractile function is depressed, the hypoperfused myocardium retains its responsiveness to an inotropic challenge.14 ,15 When, after 85–90 minutes of sustained moderate regional ischaemia (reduction of transmural blood flow by about 50%) in anaesthetised pigs, dobutamine or calcium was infused selectively into the ischaemic region, contractile function transiently increased, although regional blood flow remained reduced (fig 2). Thus an energy reserve was available in the ischaemic myocardium which was not used to maintain baseline function but could be recruited to increase contractile function transiently during an inotropic challenge. These results strongly suggest that the decrease in contractile function secondary to a reduction in myocardial blood flow was not simply the result of a reduced energy supply, but rather reflected an active adaptive process in the myocardium. Imposition of an inotropic stimulus on the short term hibernating myocardium disrupted this adaptive process, as indicated by the subsequent return to decreased myocardial creatine phosphate content and increased lactate production. A persistent inotropic reserve in response to dobutamine was also apparent in anaesthetised pigs with 24 hours of coronary stenosis and reduced resting blood flow. The inotropic response to dobutamine was typically biphasic, with increased wall thickening at lower doses and contractile dysfunction at higher doses, and was associated with increased net lactate production.20 Similarly, in dogs the response to dobutamine in collateral dependent, dysfunctional, but viable (from histology) myocardium was biphasic four weeks after implantation of ameroid constrictors.29 In both studies, recovery of contractile function following restoration of blood flow was observed. An enhancement of the inotropic response to low dose dobutamine with the addition of glyceryl trinitrate was reported in the pig model with 24 hours of coronary stenosis, suggesting that inotropic reserve is in part dependent on coronary reserve.33
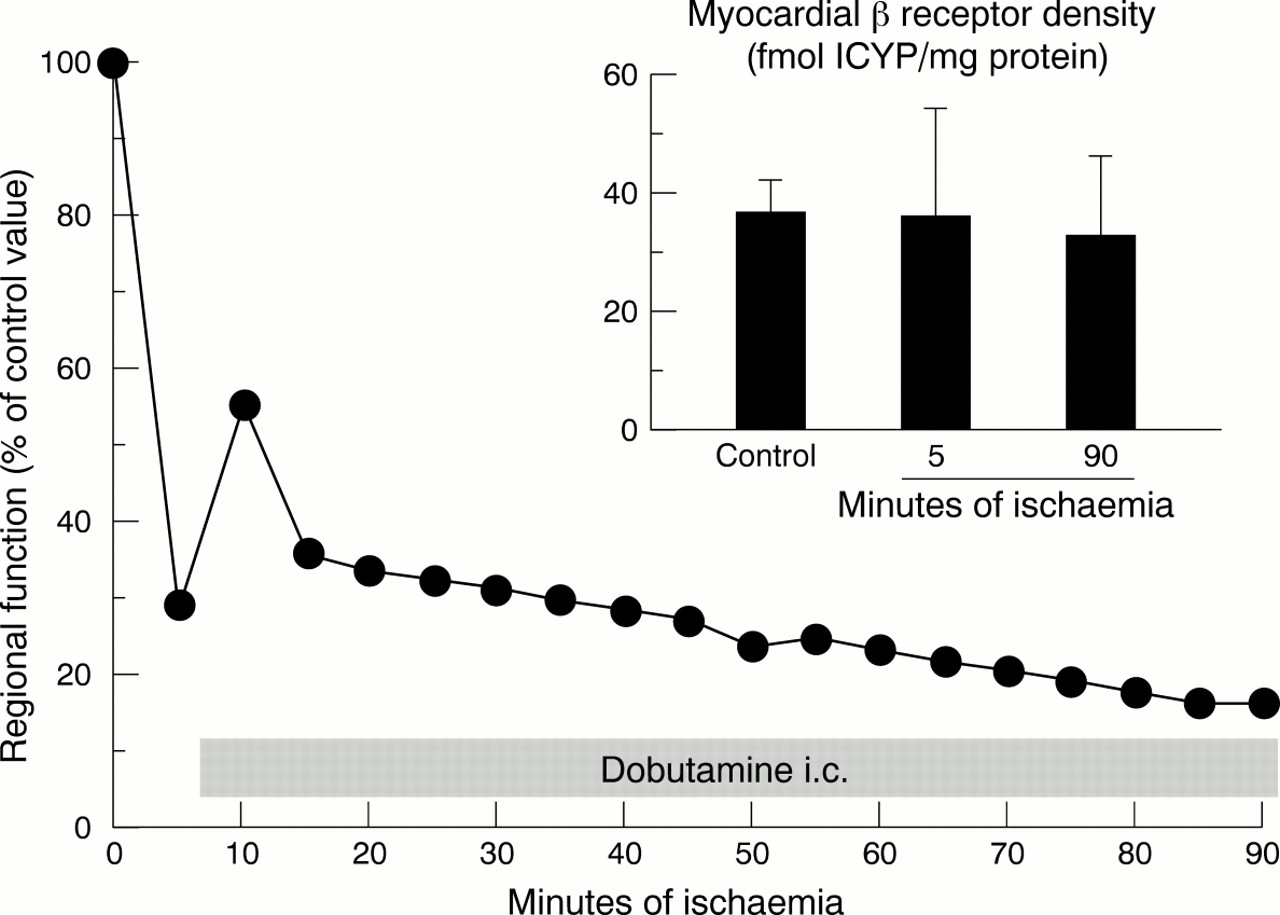
Infusion of dobutamine during moderate ischaemia results in only a transient increase in regional function. After 85 minutes of dobutamine infusion, however, regional work is reduced below the initial ischaemic value. Inset: Myocardial β adrenoceptor density does not change from control to five minutes of ischaemia and also remains constant throughout the subsequent 85 minutes of ischaemia with the additional dobutamine infusion.
Persistent inotropic reserve—that is, improved regional wall motion in response to dobutamine—has also been reported quite consistently from various studies in patients with chronic hibernation.23 ,34-36 The recruitment of an inotropic reserve was associated with increased oxidative metabolism, as evident from the increased rate constant of11C-acetate clearance.36 During recruitment of inotropic reserve with intravenous dobutamine in the dysfunctional region, glucose utilisation—as measured by18FDG positron emission tomography (PET)—was increased, whereas it was decreased in normal myocardium.23 In addition, increased anaerobic glycolysis with a significant reduction in net lactate uptake and even net production in some patients—as measured from the arteriocoronary venous differences—was seen in the dysfunctional myocardium during recruitment of an inotropic reserve with intracoronary dobutamine.34Persistent inotropic reserve, however, is also observed in chronically dysfunctional myocardium with normal perfusion35and—unless associated with metabolic deterioration—does not distinguish hibernating from stunned myocardium.
LIMITS OF SHORT TERM HIBERNATION
The development of a delicate balance between regional myocardial blood flow and function during early ischaemia is disturbed by subsequent unfavourable alterations in supply and demand. When, after five minutes of ischaemia at a blood flow reduction compatible with the development of myocardial hibernation over 90 minutes, anaesthetised pigs were subjected to a further reduction in myocardial blood flow, necrosis developed. The lower limit of transmural myocardial blood flow compatible with the development of short term myocardial hibernation over 90 minutes of coronary hypoperfusion corresponded to approximately 50% of baseline in this experimental setting, and the lower limit of subendocardial blood flow corresponded to 25% of baseline.37 Also, a further increase in energy demand by continuous inotropic stimulation with dobutamine for 85 minutes induced necrosis.37 Thus both a further reduction in energy supply by increasing severity of ischaemia and enhanced energy expenditure by continuous inotropic stimulation can impair the development of short term myocardial hibernation and precipitate myocardial infarction.
Morphology in hibernating myocardium
With five hours38 to 24 hours16 of partial coronary artery stenosis in dogs38 or pigs,16 the number of myofibrils was reduced and myofibrils were partially disrupted and aggregated around the nucleus,38 while mitochondria and glycogen deposits were increased; these alterations were reversed seven days after release of the stenosis.16 In myocardial biopsies from patients with prolonged contractile dysfunction that was reversed by bypass surgery, the amounts of thin filament complexes and titin were reduced and the remaining myofibrils, as well as the sarcoplasmic reticulum, were disorganised. Numerous small, doughnut-like mitochondria were apparent, as well as glycogen deposits.11 ,39-44 The interstitium contained cellular debris, increased numbers of macrophages and fibroblasts, and increased collagen.41 ,42 ,44-46Recently, apoptosis was also detected in human hibernating myocardium,42 and—as well as the above alterations17 ,21 ,47 ,48—was confirmed in pigs with chronic coronary stenosis.47
To a certain degree these degenerative alterations appear to be reversible.41 In addition to degenerative changes, particularly interesting changes in the distribution of titin and cardiotin have been observed, and it has been claimed that these reflect an embryonic phenotype pattern.45 It has been suggested that this, together with the expression of α smooth muscle actin in hibernating myocardium, represents hibernation induced dedifferentiation of cardiomyocytes.45 Thus it is currently unclear whether the morphological changes seen in hibernating myocardium reflect adaptive atrophy such as occurs in any quiescent muscle (which might be accompanied by dedifferentiation and an embryonic phenotype pattern) or whether the observed changes reflect degenerative, pathological, and possibly irreversible events.
Mechanism of myocardial hibernation
While β adrenoceptor density and affinity is unaltered in short term hibernating myocardium following 90 minutes of moderate ischaemia,37 the density of myocardial β adrenoceptors has been shown to be reduced in chronic animal models of myocardial hibernation29 or chronic stunning,49 as well as in patients with heart failure.50 ,51 Coronary artery bypass grafting in patients with heart failure not only improved overall ventricular function, but also increased the lymphocyte β adrenoceptor density towards normal values.52 Thus alterations in the β adrenoceptors are not responsible for the early phase of the development of myocardial hibernation, but may contribute to chronic alterations of baseline contractile function.
Activation of ATP dependent potassium channels and increases in the concentration of interstitial adenosine are not involved in the development of short term myocardial hibernation. This conclusion is based on the observation that neither the ATP dependent potassium channel blocker glibenclamide nor an increase in catabolism of adenosine caused by intracoronary infusion of adenosine deaminase altered the characteristics of short term hibernation, including perfusion–contraction matching, recovery of metabolic variables, inotropic responsiveness, and maintenance of myocardial viability.53
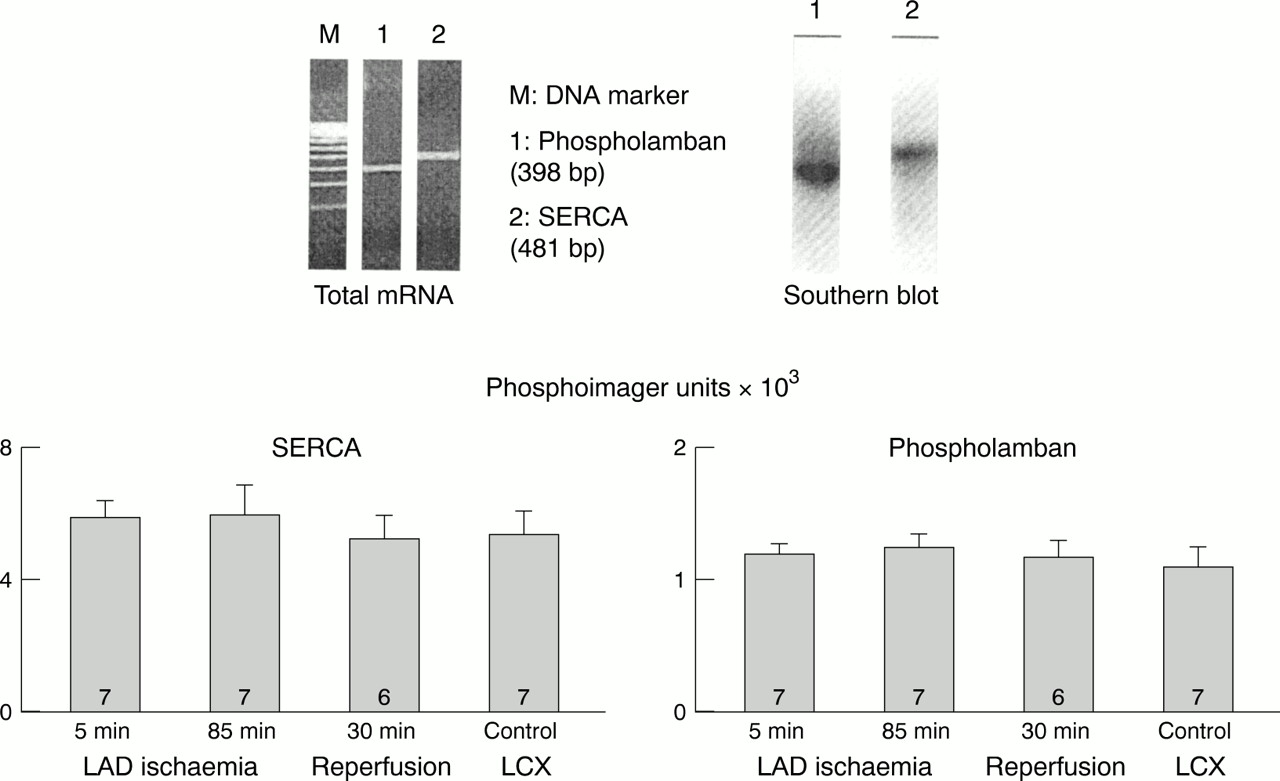
It is not surprising that, in the search for possible mechanisms of hibernation, attention has been directed to calcium. In isolated buffer perfused ferret hearts moderately decreased coronary perfusion resulted in decreased calcium transients and decreased contractile force without any change in high energy phosphates.54 Overall calcium responsiveness of short term hibernating myocardium in anaesthetised pigs is substantially reduced; however, this reduction is attributable to a decrease in maximum developed force rather than to any decrease in calcium sensitivity (fig 3).15 The expression of calcium regulatory proteins (SERCA, phospholamban, calsequestrin, troponin inhibitor) was not altered during 90 minutes of experimental short term hibernation (fig 4),55 but such alterations may contribute to contractile dysfunction over longer periods.56

(A): Increasing doses of intracoronary calcium increase regional work stepwise, both during normoperfusion and at 85 minutes of ischaemia. With ischaemia, the dose–response relation is displaced downwards. (B) The relations between the fractional increments in regional work and added intracoronary calcium are not different during normoperfusion and ischaemia. Modified from Heusch G, et al. Circulation 1996; 93:1556–66, with permission of the publisher.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of the calcium regulatory protein SERCA and phospholamban. The expression of SERCA and phospholamban is not altered during experimental short term hibernation and subsequent reperfusion. Data are means, error bars = SEM. LAD, left anterior descending coronary artery; LCX, left circumflex coronary artery. Reproduced from Lüss H, et al. Cardiovasc Res 1998;37:606–17 , with permission of the publisher.
In anaesthetised swine, endogenous nitric oxide is not involved in the immediate downregulation of baseline contractile function and the recovery of myocardial energetics, as both perfusion–contraction matching and the recovery of the free energy change of ATP hydrolysis also occurred with inhibition of nitric oxide synthesis by L-NA (NG-nitro-L-arginine). However, at any given blood flow and oxygen consumption, contractile function was lower without than with endogenous nitric oxide. Thus endogenous nitric oxide does not appear to mediate the immediate downregulation of baseline contractile function but to contribute to successful hibernation by reducing futile oxygen consumption and maintaining contractile function at the best possible level without any additional energetic costs, most probably through preservation of contractile calcium sensitivity.57
Thus, to date, the mechanisms responsible for the development and maintenance of hibernation remain unclear.
Blood flow in myocardial hibernation: persistent ischaemia or cumulative stunning, or both?
As mentioned above, a series of acute experimental studies with controlled coronary hypoperfusion identifies features consistent with myocardial hibernation—that is, perfusion–contraction matching, recovery of myocardial substrate and energy metabolism during ongoing ischaemia, persistent inotropic reserve, and lack of necrosis (for review, see Heusch10). Bolli58 was the first to propose that the phenotype of hibernating myocardium—chronic yet reversible contractile dysfunction in the setting of coronary artery disease—could also arise from repetitive or cumulative stunning with or without (perfusion–contraction mismatch) an underlying reduction in baseline blood flow. In most chronic animal experiments and patient studies, myocardial blood flow and function have been measured at only a single time point, and the potentially diseased areas have been compared with those identified as normal.59 Based on such single measurements, animals or patients were grouped according to their flow–function relation as either having hibernating myocardium (reduced flow and reduced function—that is, perfusion–contraction matching) or stunned myocardium (normal or almost normal flow and reduced function—that is, perfusion–contraction mismatch). It is only recently that several acute and chronic animal experiments have focused on studying the temporal progression towards myocardial hibernation by measuring flow and function at several time points. In anaesthetised pigs with controlled coronary hypoperfusion, perfusion–contraction matching was initially maintained only for 90 minutes; with prolongation of ischaemia to 24 hours, contractile function was further reduced, despite an unchanged regional myocardial blood flow.60 Apparently, with ischaemia of longer than 90 minutes' duration, additional factors acted to reduce contractile function, thereby inducing a perfusion–contraction mismatch. However, the close matching between myocardial blood flow and oxygen consumption was maintained, which continues to support the concept of metabolic adaptation to prolonged ischaemia.60
Several chronic animal studies (reviewed by Canty and Fallavollita61) show an early period in which contractile dysfunction is associated with normal or almost normal resting perfusion but reduced coronary reserve (consistent with the idea of chronic repetitive stunning), while with time and the progression of contractile dysfunction, resting myocardial blood flow is also reduced (consistent with the idea of hibernating myocardium). Furthermore, within a given heart in chronically instrumented animals62or in patients,63 hibernation and stunning might coexist, as some (32–50%) but not all areas with contractile dysfunction have reduced myocardial blood flow.
Thus in our view the distinction between reduced baseline blood flow and repetitive stunning is artificial when based on single point measurements, and distracts from the concept of myocardial hibernation. As originally proposed by Bolli,58 many patients with coronary artery disease will have both reduced baseline flow and superimposed repetitive episodes of stress induced ischaemia with subsequent stunning in their daily life. However, patients suffering from acute myocardial ischaemia will either adapt to the state of hypoperfusion—that is, develop a state of short term myocardial hibernation—or otherwise develop extensive myocardial infarction. As many patients, especially in the scenario of unstable angina, undergo acute episodes of myocardial ischaemia, short term myocardial hibernation may be a much more common and important phenomenon than chronic myocardial hibernation.64
Clinical importance of hibernation
Not surprisingly, data on the incidence of hibernating myocardium are scanty in patients with coronary artery disease. Established diagnostic tests for the identification of hibernating myocardium include the demonstration of a mismatch between reduced flow and enhanced glucose uptake using PET techniques65-68 and dobutamine echocardiography.69 A positive low dose dobutamine test is a good predictor of functional recovery on revascularisation70-74 and a good predictor of survival.75 ,76 Dobutamine echocardiography has a higher false negative rate than PET,77 and a negative dobutamine test does not exclude hibernation, as the associated loss of myofibrils will also induce a loss of inotropic reserve.35 ,78Contractile reserve during dobutamine echocardiography also correlates inversely with the extent of interstitial fibrosis.79 ,80Other tests for hibernating myocardium are based on the uptake and retention of nuclear tracers such as thallium and sestamibi.74 ,81-83 Sestamibi scintigraphy and histological evidence of viability correlate well (r = 0.89).84
Clinical syndromes consistent with the existence of myocardial hibernation include unstable and stable angina,11 ,85-89acute myocardial infarction,1 ,85 ,90-92 left ventricular dysfunction with or without congestive heart failure,93 ,94 and the ALCAPA syndrome (anomalous left coronary artery from the pulmonary artery).95-97
In patients studied 5–21 days after an acute myocardial infarct, 78% had perfusion–metabolism mismatch in at least one myocardial area on PET scanning, indicating hibernating myocardium.92Likewise, 69% of patients with an acute myocardial infarction had a further reduction in the perfusion deficit using sestamibi tomography between five weeks and seven months after the infarction, associated with improved wall motion and suggesting hibernating myocardium.98 In patients with a left ventricular ejection fraction below 30% because of ischaemic heart disease, 36%11 or 52%99 had significant areas of hibernating myocardium as identified by PET. Patients with a left ventricular ejection fraction < 30% benefit more from coronary revascularisation than those with an ejection fraction > 30%.100 It has been suggested that as many as 11% of the patients referred for heart transplantation have hibernating myocardium.94 ,101
The available data on the prevalence of hibernation vary substantially. It is possible that our awareness of the possible existence of the phenomenon as well as the eventual availability of simple standard methods for its identification will prove that hibernation is more common than currently recognised. Certainly, at the present time it is easier to miss it than to find it!
Conclusions
The myocardium can adapt to prolonged moderate ischaemia for several hours. Such adaptation is associated with recovery of contractile function during reperfusion and lack of necrosis. Reduced blood flow at rest is also observed in dysfunctional regions of conscious, chronically instrumented animals with chronic coronary stenosis, consistent with the original concept of Rahimtoola.4 Also, morphological alterations in such animal models with chronic coronary stenosis are remarkably similar to those found in patients with chronic hibernation. The existence of cumulative stunning secondary to repeated episodes of stress induced ischaemia in regions with normal blood flow at rest but with reduced coronary reserve was proposed as an alternative mechanism underlying the observed regional contractile dysfunction in studies with chronic coronary stenosis. This possibility certainly exists—as do other more complex scenarios of flow and function in the setting of coronary stenosis58—but has not been systematically investigated and, above all, does not exclude the existence of hibernation in other studies. In patients, the majority of quantitative data available, however, suggest that there is reduced blood flow at rest in the dysfunctional region compared with remote reference regions in the same individual.
Acknowledgments
The authors' studies were supported by the Deutsche Forschungsgemeinschaft, the IFORES program of the Medical School of the University of Essen, the German Cardiac Society, and the Hans und Gerti Fischer Stiftung.